Diez Group
Our interdisciplinary lab is interested in the development and the application of optical techniques to investigate molecular transport in cell biology and nanotechnology.



 © Diez
© Diez
Molecular Transport in Cell Biology and Nanotechnology
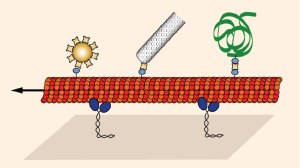
Using methods of single-molecule biophysics and in vitro reconstruction we study cooperative effects in motor transport and cell motility. Moreover, we apply molecular motors towards the synthesis of nanomaterials, molecular diagnostics, surface imaging and parallel biocomputation. Read more about our research.




Prof. Dr. Stefan Diez
Send encrypted mail via the SecureMail portal (for TUD external users only).